
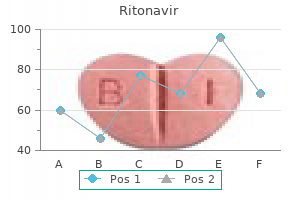
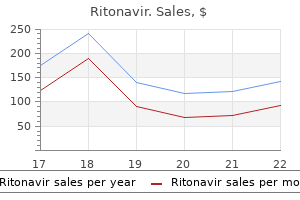
"Cheap 250mg ritonavir visa, medicine park lodging".
K. Daro, M.A., M.D.
Co-Director, University of Arizona College of Medicine – Tucson
The nettosuchids had a unique jaw articulation and typically a broad, elongate snout. Melanosuchus and Caiman appear only in the Neotropic Late Miocene and Pleistocene, respectively. In contrast, Alligator ranges from the Early Oligocene to the present in North America and Asia. Crocodyloids similarly had a moderate diversity in the Late Cretaceous and Early Tertiary. The tomostomines occurred in the Middle Eocene of Egypt and China, then intermittently in northern Africa and Europe from the Oligocene to the Middle Miocene and then not again until the Late Pliocene in Asia. The crocodylines include a variety of lineages of which the "true" Crocodylus is of only recent origin from the Pliocene to the present. The mekosuchines had a variety of body and head forms, ranging from narrow elongate skulls like gharials to short, broad-headed species. Mekosuchus survived into the Recent era in New Caledonia and apparently was hunted to extinction by the first humans to arrive there. The ribs and vertebrae were fused to dermal bones to form a carapace, and some pectoral girdle elements and dermal bones fused to form a plastron. A pleurodire, Proterochersis, was contemporaneous and sympatric in Europe with Proganochelys. The pelvic girdle was fused to the plastron, indicating that it was the earliest pleurodire and confirming that the divergence of cryptodires and pleurodires had occurred. After Proterochersis, the pleurodire Platychelys occurred through much of the Jurassic and into the Early Cretaceous. By the mid-Cretaceous, pleurodire are represented in many fossil faunas, particularly those of the Southern Hemisphere. Although now confined to the southern continents, a few pleurodires occurred in the Northern Hemisphere at least through the Miocene. Some Tertiary pleurodires were marine or estuarine and reached the size of modern sea turtles, although they did not develop the morphology and locomotor mode of the cryptodiran sea turtles. Chelids do not appear until the Oligocene or Miocene and only in South America and Australia. In contrast, the fossil history of the extant pelomedusoids begins in the Early Cretaceous. Pelomedusid sidenecks occur first in the Late Cretaceous with all subsequent fossils confined to Africa. The largest turtle recorded is the Miocene podocnemidid Stupendemys geographicus, which had a carapace length of 3 meters. The oldest turtle in North America and the first cryptodire is Kayentachelys aprix, from the late Early Jurassic (185 Ma) of western North America. Thereafter, fossil cryptodires are absent until the appearance of the Pleisochelyidae and Pleurosternidae in the lower Late Jurassic; subsequently, cryptodires remained part of the reptilian fauna. Both fossil families contained moderate-sized, aquatic turtles, and neither is related to any of the later-appearing turtle groups. The pleisochelyids are structurally more advanced turtles and the sister group to the meiolaniids and all extant groups of cryptodires.
In the mid-twentieth century, the theory of continental drift was superseded and replaced by the theory of plate tectonics, which provided a clear mechanism accounting for the drifting of continents. Although we will not detail the theory here, the basic elements of continental drift are that continents ride on massive plates that slowly drift across the surface of the Earth. This surface, the lithosphere, consists of seven large tectonic plates as well as many smaller ones and is relatively viscous. Boundaries of the drifting plates converge, diverge, or transform, depending on forces underneath the plates. Convergence occurs when two plates come together, often resulting in production of mountain ranges. Subduction is a term used to describe the process by which one of the plates moves under the other. Earthquakes and volcanic eruptions are instantaneous (geologically speaking) results of major plate movements, and major changes in the distribution of landmasses are long-term results of plate movements. During the history of the Earth, landmasses have coalesced and drifted apart many times. Because the evolution of life was occurring as continents moved, studies of biogeography changed radically when continental drift and plate tectonics were accepted by the scientific community. Land bridges were no longer the only available explanation for global patterns of distribution. Land bridges of course were involved in some colonizations and recolonizations, but they were no longer the only explanations for patterns of distribution. Prior to the origin of ancestors of amphibians and reptiles (approximately 350 Ma), two large continents (Gondwana and Old Red Sandstone continent) existed. Old Red Sandstone continent combined with several smaller continents to form Laurussia, which drifted toward Gondwana. By approximately 250 Ma, a single supercontinent, Pangaea, existed, which resulted from the collision of Laurussia and Gondwana. Through time, Pangaea split into two large continental masses, Gondwana and Laurasia, each of which split further into the continents that we see today. Although patterns of distribution have interested scientists for at least 200 years, biogeography studies prior to about 1967 were largely descriptive. Wilson published their classic book the Theory of Island Biogeography in 1967, the field changed dramatically, and biogeography became a predictive rather than a descriptive science. We already introduced some elements of island biogeography in the last chapter, in an ecological context. To reiterate the key points, the theory of island biogeography posits that the number of species in a given area can be predicted based on a few key variables: island size, immigration rate, and extinction rate. Most studies of island biogeography reveal a close association between the number of species and island size, if all else is equal. Distance of islands from mainland habitats (sources of species) also affects number of species (richness). The farther an island is from the mainland, the fewer species are likely to be present, and this variation is largely a result of reduced immigration rates or increased extinction rates. Island biogeography has much wider application because habitats distributed across the Earth are patchy, and each patch can effectively be considered an island. Thus the underlying hypotheses associated with the theory of island biogeography can be tested in studies of aquatic, marine, and terrestrial organisms. The theory of island biogeography can be applied to ecological questions, particularly in landscape ecology and conservation biology. Unlike plate tectonics, land bridges, and changes in ocean levels, island biogeography centers on patch size and distance from sources and is most applicable to predicting numbers of species and immigration and extinction rates in a relatively short time period (ecological time). Historical biogeography (plate tectonics Chapter 13 Biogeography and Phylogeography 383 and its correlates) centers on long-term (geologic time) patterns of distribution and diversification and thus correlates origins and patterns of diversification of faunas with the geologic history of the Earth. Late in the twentieth century, with the rapid development of molecular systematics, historical biogeography was, for all practical purposes, reinvented. The potential existed to estimate time periods during which major evolutionary shifts occurred in a particular group of organisms based on calibrated molecular clocks and then fit patterns of divergence to independently derived estimates of time based on the fossil record and geological data.
Ecology of the elusive tropical lizard Tropidurus [=Uracentron] flaviceps (Tropiduridae) in lowland rain forest of Ecuador. Ecology of the nocturnal lizard Thecadactylus rapicauda (Sauria: Gekkonidae) in the Amazon region. Observations on the ecology of the rare Amazonian lizard, Enyalius leechii (Polychrotidae). The impact of individual tree harvesting on thermal environments of lizards in Amazonian rain forest. Pushing the edge: Extended activity as an alternative to risky body temperatures in a herbivorous teiid lizard (Cnemidophorus murinus: Squamata). Detecting variation in microhabitat use in low-diversity lizard assemblages across small-scale habitat gradients. Use of time, space, and food by the gymnophthlamid lizard Prionodactylus eigenmanni from the western Amazon of Brazil. Small in a big world: Ecology of leaf-litter geckos in new world tropical forests. Ecological variation among populations of the gekkonid lizard Gonatodes humeralis in the Amazon Basin. Ecology of the lizard Norops oxylophus (Polychrotidae) in lowland forest of southeastern Nicaragua. Ecology of the lizard Kentropyx pelviceps (Sauria: Teiidae) in lowland forest of Ecuador. Reproductive behaviour of the glass frog Hyalinobatrachium valerioi (Anura: Centrolenidae) at the tropical stream Quebrada Negra (La Gamba, Costa Rica). Effects of incubation temperature on sex determination in a community of Neotropical freshwater turtles in southern Mexico. Toward the origin of amniotes: Diadectomorph and synapsid footprints from the early Late Carboniferous of Germany. Structure and seasonal variation of the male reproductive organs of Vipera berus (L. Habitat utilization, movements, and activity patterns of Enhydris plumbea (Serpentes: Homalopsinae) in a rice paddy wetland in Borneo. Feeding strategies in marine snakes: An analysis of evolutionary, morphological, behavioral and ecological relationships. Description and systematics of panderichthyid fishes with comments on their relationship to tetrapods. Taxonomic status of the West African Agama (Sauria: Agamidae) with prominent tail crests: Agama boulengeri Lataste 1886, Agama insularis Chabanaud, 1918 and Agama cristata Mocquard, 1905. New genera and a new species of Central American salamanders with a review of the tropical genera (Amphibia, Caudata, Plethodontidae). Direct development in the lungless salamanders: What are the consequences for developmental biology, evolution and phylogenesis The reproductive biology of Eleutherodactylus jasperi (Amphibia, Anura, Leptodactylidae), with comments on the evolution of live-bearing systems. Reproduction, growth, and population structure of the Central American caecilian Dermophis mexicanus. Non-traditional characters in the assessment of caecilian phylogenetic relationships. Cartilage in the cloaca: Phallodeal spicules in caecilians (Amphibia: Gymnophiona). A brief history of research on gymnophionian reproductive biology and development. Visceral anatomy of the Malaysian snake genus Xenophidion, including a cladistic analysis and allocation of a new family (Serpentes: Xenophidiidae).
Syndromes
- Lack of tears
- You may feel dizzy and off-balance for anywhere from 20 minutes to 24 hours.
- Death among children and adolescents
- Echocardiogram
- Cornelia de Lange syndrome
- Born to a mother with diabetes
- Heart attack, or any disease of the heart that weakens or stiffens the heart muscle (cardiomyopathy)
- Sore throat
- Megaloblastic anemia
As with sea turtles, the flipper-shaped forelimbs are the major locomotor appendages and propel the animal using a figure-eight stroke. This type of stroke mimics underwater flying in sea turtles and penguins and apparently is used predominantly for slow and moderate-speed locomotion; when pursued, the turtle reverts to the typical quadrapedal swimming gait of other aquatic turtles. Nesting occurs in the latter part of the dry season, mainly from August to October when the river sandbanks and bars are exposed. Trionychidae Softshell Turtles Classification: Reptilia; Eureptilia; Testudines; Cryptodira. Clockwise from upper left: Pig-nose turtle Carettochelys insculpta, Carettochelyidae (R. Characteristics: Softshells are flattened, pancake-shaped turtles that have reduced bony carapaces and plastrons. The carapace and plastron are naked, lacking epidermal scutes, but are covered with a thick, leathery skin. The flattened carapace lacks peripheral bones (except in Lissemys), and the nuchal lacks costiform processes. The neck withdraws vertically; this mechanism is reflected in an anteroventrally oriented articular surface of the first thoracic vertebra. Other vertebral traits are the exclusion of the 10th thoracic vertebra from the sacral complex and procoelous caudal vertebrae. Content: Three genera, Cyclanorbis, Cycloderma, and Lissemys, each with 2 species. Characteristics: the lattice-like plastral skeleton has bilaterally fused hyoplastral and hypoplastral bones, and externally the plastron has well-developed femoral flaps. The biology of the African taxa Cycanorbis and Cycloderma is little studied; the South Asian Lissemys is somewhat better known. They actively forage and also lie partially hidden in the bottom silt or sand, waiting for passing prey. They are presumably opportunistic omnivores, eating invertebrates, small vertebrates, and occasional plant matter. Content: Ten genera, Amyda, Apalone, Chitra, Dogania, Nilssonia (including Aspideretes), Palea, Pelochelys, Pelodiscus, Rafetus, and Trionyx, with 1, 3, 3, 1, 5, 1, 3, 4, 2, and 1 species, respectively. Distribution: Eastern North America, South Asia to Japan southward to New Guinea, and north-central sub-Saharan Africa into Southwest Asia. Characteristics: the lattice-like plastral skeleton has separate hyoplastral and hypoplastral bones on each side, and externally the plastron lacks femoral flaps. All are highly aquatic turtles, spending much of their time partially buried on the bottom waiting for prey. Their long necks and protruding, snorkel-like snouts permit them to extend their noses to the water surface to breathe; they also depend to some extent upon cutaneous respiration. They actively forage for prey, and their flattened hydrodynamically efficient habitus makes them excellent and fast swimmers. Dogania subplana appears to be the only softshell that occurs in small mountain streams. All trionychines are predominantly carnivorous, although they likely feed on plant matter, particularly when animal prey is not readily available. Temperate and subtemperate species are predominantly spring breeders, and tropical species lay eggs in the early dry season. Incubation generally requires 8 to 10 weeks, although in Nilssonia gangeticus, it is 36 to 42 weeks or as brief as 28 days in Pelodiscus sinensis. Cheloniidae Hard-Shelled Sea Turtles Classification: Reptilia; Eureptilia; Testudines; Cryptodira. Content: Five genera, Caretta, Chelonia, Eretmochelys, Lepidochelys, and Natator, with 1, 1, 1, 2, and 1 species, respectively. They have flattened, streamlined shells covered with epidermal scutes and forelimbs modified into large flippers.